The mechanism of the action potential. Department of Physiology Membrane action potential of its phase
Biological reaction - the response of cells, tissues, organs in response to an irritant (stimulus).
Irritability- the property of all living tissues to change their internal state when external conditions change.
Types of tissues depending on the response to external stimuli:
I Excitable- have the property of excitability, i.e. the ability to be excited nervous, muscular, glandular.
II Non-excitable- change their state, but do not generate an excitation process in response to the applied stimulus.
Excitability- the ability of the tissue to go into an excited state.
Excitation- the active state of tissues in response to the action of an irritant, this is a complex biological reaction, manifested in the totality of physical, physico-chemical and functional changes, capable of spreading through the tissue.
Excitation includes non-specific and specific components.
Non-specific:
shift of chemical reactions, generation of heat, physical and chemical changes,
production of biopotentials, structural changes in the cell membrane.
Specific:
muscle tissue responds with muscle contraction, nervous tissue - with the generation of a nerve impulse and its conduction, glandular tissue - with the formation and secretion of a secret.
Excitation can be local and dynamic (propagating).
Biopotentials
Luigi Galvani in 1791 in an experiment showed that living tissues contain "animal electricity", his scientific opponent, the physicist Volta - that this is electricity from dissimilar metals, he created the first direct current source, which is called a galvanic cell.
Types of biopotentials:
1. Resting biopotential (membrane) - MPP.
2. Biopotential of action (excitation) - PD.
- resting biopotential is the potential difference between the outer and inner surface of the cell membrane at rest. The outer surface of the cell membrane is positively charged, while the inner surface is negatively charged.
The resting biopotential is recorded by the intracellular method - using microelectrodes, one of which is inserted into the cell (Fig. 1).
Figure 1. Schematic representation of the method for recording biopotentials.
In the experiment, the resting biopotential can be registered between the damaged and undamaged tissue area. The damaged area is a model of the inner surface of the cell membrane.
During intracellular recording, membrane recharging is recorded under one electrode (single-phase AP), while during extracellular recording, the action potential passes through two electrodes (two-phase AP is recorded).
- Biopotential action- these are short-term high-amplitude changes in the MPP that occur during excitation. AP is recorded in irritated tissues, in which an excitation wave occurs (Fig. 2). PD is measured using an intracellular lead and an extracellular lead.

Figure 2. Action potential, its main phases.
Modern, experimentally proven, membrane-ionic theory of the origin of biopotentials (Hodgkin, Huxley, Katz).
Basic provisions:
- Electrical processes occur on the plasma membrane of the cell, which consists of a bimolecular layer of lipids (the backbone of the membrane) and proteins that perform various functions in the membrane: receptor, enzymatic, form channels and pumps in it (Fig. 3).
The membrane channel can be non-specific, it is constantly open, does not have a gate mechanism, and electrical influences do not change its state. It is called a "leak" channel. Specific channels (selective) have a gate mechanism, so they can be either open or closed, depending on the electrical effects on the membrane, and allow only a certain ion to pass through. This channel consists of three parts: water pore - lined inside with hydrophilic groups; selective filter - on the outer surface, which passes ions depending on their size and shape; gate - on the inner surface of the membrane, control the permeability of the channel.

Figure 3. Structure of a biological membrane.
Membrane channel may be non-specific, it is constantly open, does not have a gate mechanism, electrical influences do not change its state. It is called a "leak" channel. Specific channels (selective) have a gate mechanism, so they can be either open or closed, depending on the electrical effects on the membrane, and allow only a certain ion to pass through. This channel consists of three parts: water pore - lined inside with hydrophilic groups; selective filter - on the outer surface, which passes ions depending on their size and shape; gate - on the inner surface of the membrane, control the permeability of the channel (Fig. 4).

Figure 4. The structure of the ion channel.
Channels for sodium have two types of gates: fast activation and slow inactivation. At rest, slow inactivation pathways are open and fast activation pathways are closed. Upon excitation, fast activation ones open and slow inactivation ones close, i.e. for a short period of time, both types of gates are open (Fig. 5).

Figure 5. Operation of the activation and inactivation gate mechanisms of the sodium ion channel.
potassium channels have only slow gates.
Pumps carry out the function of transporting ions across the membrane against a concentration gradient; ATP energy is used for their operation.
- There is a concentration gradient on both sides of the membrane.
Inside the cell 40 times > K+; t;/p>
Outside the cell: 20-30 times > Na+,
50 times > Cl-.
- The membrane allows molecules of fat-soluble substances to pass through, but anions of organic acids do not. The membrane is permeable to water; for ions, the permeability of the membrane is different: for potassium at rest, the permeability is almost 25 times greater than for sodium. When excited, the permeability to both potassium (gradually) and sodium (quickly, but for a very short period of time) increases.
resting potential
The permeability of the membrane for K+ ions is increased, so potassium plays a major role in the generation of MPP. Potassium creates an electric field and charges the outer surface of the "+" membrane. At that moment, when the “+” potential of the outer side reaches a certain value in relation to the “–” inside, which is created by anions, a dynamic balance occurs between the K+ ions entering and exiting the cell. This moment corresponds to the equilibrium potential for K - the resting potential.
MPP is characterized by:
1. constancy;
2. polarity, outside "+", inside "-";
3. value - in mV, for skeletal muscle - 60 - 90 mV,
for smooth - -30 - 70 mV,
for the nerve -50 - 80mV,
for a secretory cell - -20mV.
MPP is one of the main indicators of the state of physiological dormancy of the cell. With an increase in the extracellular concentration of potassium, the MPP decreases, because. the diffusion of potassium from the cell decreases due to a decrease in its concentration gradient. Under the action of substances that block the resynthesis of ATP, tk. the work of the sodium-potassium pump stops, the MPP also decreases. Sodium and chloride ions enter the cell, but due to the low permeability, they do not have a significant effect on the magnetic field.
action potential
When excited, the permeability for Na ions increases sharply (several thousand times), which enter the cell like an avalanche and charge the inner side of the “+” - the membrane depolarizes, and then the number of sodium ions inside exceeds the potassium charge on the surface and this leads to a recharge of the membrane (reversions). Gradually increasing permeability to potassium and its flow out of the cell inactivates sodium permeability and leads to restoration of the charge on the membrane. There is a repolarization phase.
A significant factor is the sodium-potassium pump, which removes 3 sodium ions from the cell in exchange for 2 potassium ions introduced into the cell. Its work depends on the metabolism of the cell, in particular, on its energy supply. In this case, 1 ATP molecule is consumed (Fig. 6).

Figure 6. Mechanism of the sodium-potassium pump.
AP consists of a peak potential, which is formed by the phase of depolarization, reversion and repolarization, and trace potentials (Fig. 2).
Trace potentials:
Negative (trace depolarization);
Positive (trace hyperpolarization).
The reason for trace potentials is further changes in the ratio between the entry of sodium into the cell and the exit of potassium from it. With trace depolarization, a residual current of sodium into the cell is noted, while a decrease in potassium current is noted. With trace hyperpolarization - a residual increase in the current of potassium from the cell with simultaneous activation of the sodium-potassium pump.
PD is characterized by:
1. changing character;
2. short duration - a few milliseconds;
3. membrane charge, outside - "-", inside - "+".
Under the action of substances that block sodium channels, PD is not generated, because. Normally, membrane depolarization is due to an increase in its sodium permeability. With an increase in the strength of the stimulus above the threshold, the AP amplitude does not change, because the number of activated sodium channels does not change, which open to the maximum at threshold stimulation.
Conditions necessary for the occurrence of excitation (laws of irritation).
Excitability of tissues is different. To cause arousal, the stimulus must have:
1. By sufficient force - the law of the threshold.
2. The steepness (gradient) of the growth of this force is the law of accommodation.
3. Time of action - the law of force-time.
1. The law of strength. A measure of excitability is the threshold of irritation - the minimum strength of the stimulus that can cause excitation. All stimuli can be divided into subthreshold, threshold and suprathreshold. According to their biological significance, stimuli are divided into adequate (acting on the tissue in natural conditions, it is adapted to them in the process of evolution) and inadequate. In physiological experiments, an electric current is most often used as a stimulus, because. it causes reversible changes, is easily dosed in strength and duration, and by its nature is close to electrical processes occurring in living organisms.
In 1870, in an experiment on the heart muscle, by applying single threshold stimuli to it, he recorded a response - he found that there was no reaction to subthreshold irritation, with threshold strength and suprathreshold amplitude of the response was the same. Based on this, he proposed the "All or Nothing" law.
After the introduction of microelectronic technology into experimental studies, it was found that a response occurs in the tissue to subthreshold irritation. If the stimulus strength is less than 50% of the threshold value, then passive depolarization occurs under the poles of the electrodes without changing the membrane permeability for ions (electrotonic changes). If the stimulus strength is less than the threshold value, but more than 50% of it, then a local response occurs in the tissue, which is accompanied by membrane depolarization in the area of irritation and does not extend to the entire tissue, the excitability of tissues in this area is increased. The local response obeys the law of force relations, i.e. the greater the strength of the subthreshold stimulus, the greater the amplitude of the local response. The permeability of the cell membrane in this area increases for sodium ions. When a threshold stimulus is applied, AP arises, the amplitude of which does not change if the stimulus value exceeds the threshold value, i.e. corresponds to the “All or nothing” law, but the duration of AP for superthreshold stimuli will be shorter due to the shortening of the duration of the local response.
The moment of transition of the local response to AP is called the critical level of depolarization (CDL), and the shift of the membrane charge from the membrane potential to the CCD is called the threshold potential, along with the threshold of irritation, it characterizes the excitability of the tissue.
Change in excitability of tissues during excitation.
When excited, the excitability of tissues undergoes certain changes depending on the phases of AP (Fig. 7):
I - supernormal excitability (primary) corresponds to a local response, while two subthreshold stimuli applied with a time interval shorter than the duration of the local response can be summed up and cause AP;
II - absolute refractoriness - corresponds to regenerative depolarization and reversion, while the tissue becomes absolutely unexcitable and does not respond to the strongest stimuli;
III - relative refractory phase, corresponds to repolarization, while the excitability of the tissue is gradually restored and the suprathreshold stimulus applied during this period can generate AP;
IV - supernormal excitability (secondary or exalphatic phase) - trace depolarization, the tissue becomes more excitable than in the initial state, and even a subthreshold stimulus can cause AP;
V - subnormal excitability - trace hyperpolarization, tissue excitability is somewhat reduced.

Figure 7. Changes in membrane excitability during the development of an action potential.
2. The excitation gradient law (Dubois Reymond). The greater the stimulation gradient, the greater (up to certain limits) the reaction of the living formation.
During the action of a slowly increasing stimulus, tissue adaptation occurs - accommodation. It is due to the fact that during excitation, the permeability for sodium ions increases for a short period of time, if during its stimulus the stimulus does not reach the threshold value, then the increasing permeability for potassium ions inactivates sodium permeability and excitation does not occur. In this case, the FRA also shifts with an increase in the threshold potential.
3. The law of force-time (Lapik). The threshold value of any stimulus is inversely related to the time of its action, which is characterized by a mathematical curve - a hyperbole. The nature of the curve indicates that subthreshold stimuli (less than 1 rheobase) will not cause excitation no matter how long they act, at the same time, a very strong short-term stimulus, the duration of which is less than the useful time, will also not cause excitation.
The direct current force, which, acting indefinitely, causes excitation, is called rheobase.
The time during which the current in 1 rheobase causes excitation is useful time.
The minimum time during which a current of 2 rheobases will cause excitation is called chronaxy. The study of this indicator is used in neurological and traumatological practice to study the dynamics of recovery in the nervous or muscle tissue after an injury.
List of used literature
- Normal physiology / Ed. V.I. Filimonova. - K. - Health, 1994. - S. 5 - 37.
- Human Physiology / Ed. G. I. Kositsky. - M., Medicine, 1985. - S. 19 - 84.
- Posіbnik s normal fіzіologiї / Pіd ed. V.G. Shevchuk. - K., Health, 1995. - S. 6 - 36.
- Guide to practical exercises in physiology / Ed. G. I. Kositsky. - M., Medicine, 1988. - S. 72 - 94.
- Normal Physiology / Ed. V. I. Filimonova. - Zaporozhye, 1995. - S. 74-72.
- Human physiology. T.1 / Ed. R. Schmidt and G. Thevs. - M., Mir, - 1996. - S. 9-87.
- Human physiology. T.1 / Ed. V.M. Pokrovsky. - M., Medicine, 1998. - S. 27-97.
- General course of human and animal physiology. T.1. / Ed. HELL. Nozdrachev - M., Higher School, 1991.- P.36-116.
- Human physiology. / Ed. V.M. Smirnova - M., Medicine, 2002. - S. 45-61, 82-94.
- Physiology of people. William F. Ganong. - BaK, Lviv, 2002. - S. 6 - 69, 74-76.
An action potential is a rapid change in the membrane potential that occurs when nerve, muscle, and some glandular cells are excited. Its occurrence is based on changes in the ionic permeability of the membrane. In the development of the action potential, four successive periods are distinguished: 1) local response; 2) depolarization; 3) repolarization and 4) trace potentials (Fig. 2.11).
Local response is an active local depolarization resulting from an increase in the sodium permeability of the cell membrane. The decrease in membrane potential is called depolarization. However, with a subthreshold stimulus, the initial increase in sodium permeability is not large enough to cause rapid membrane depolarization. A local response occurs not only at subthreshold, but also at suprathreshold
Rice. 2.11.
1 - local response; 2 - depolarization phase; 3 - phase of repolarization; 4 - negative trace potential; 5 - positive (hyperpolarization) trace potential
stimulus and is an integral component of the action potential. Thus, the local response is the initial and universal form of tissue response to stimuli of various strengths. The biological meaning of the local response is that if the stimulus is small in strength, then the tissue reacts to it with a minimum expenditure of energy, not including the mechanisms of specific activity. In the same case, when the irritation is suprathreshold, the local response turns into an action potential. The period from the onset of irritation to the onset of the depolarization phase, when the local response, increasing, reduces the membrane potential to a critical level (KUD), is called the latent or latent period, the duration of which depends on the strength of the stimulation (Fig. 2.12).
Depolarization phase characterized by a rapid decrease in the membrane potential and even recharging of the membrane: its inner part becomes positively charged for some time, and the outer part becomes negatively charged. The change in the sign of the charge on the membrane is called perversion - reversion of potential. Unlike a local response, the rate and magnitude of depolarization do not depend on the strength of the stimulus. The duration of the depolarization phase in the frog nerve fiber is about 0.2-0.5 ms.
Duration phases of repolarization is 0.5-0.8 ms. The restoration of the initial value of the membrane polarization is called repolarization. During this time, the membrane potential

Rice. 2.12. Action potentials arising in response to threshold stimulation by short (A) and long-term (B) stimuli Annoying stimuli, under the influence of which responses A and B were obtained: PP - resting potential; Ekud. - critical level of membrane depolarization (according to A.L. Katalymov)
cial is gradually restored and reaches 75-85% of the resting potential. In the literature, the second and third periods are often called action potential peak.
The fluctuations in the membrane potential following the peak of the action potential are called trace potentials. There are two types of trace potentials - trace depolarization and trace hyperpolarization, which correspond to the fourth and fifth phases of the action potential. The trace depolarization (negative trace potential) is a continuation of the repolarization phase and is characterized by a slower (compared to the repolarization phase) recovery of the resting potential. The trace depolarization turns into trace hyperpolarization (positive trace potential), which is a temporary increase in the membrane potential above the initial level. An increase in membrane potential is called hyperpolarization. In myelinated nerve fibers, trace potentials are more complex: trace depolarization can turn into trace hyperpolarization, then sometimes a new depolarization occurs, only after that the resting potential is fully restored.
The ionic mechanism of the action potential. The action potential is based on changes in the ionic permeability of the cell membrane that develop sequentially over time.
When an irritant acts on a cell, the permeability of the membrane for Na + ions increases sharply due to the activation (opening) of sodium channels.
At the same time, Na + ions intensively move along the concentration gradient from outside to the intracellular space. The entry of Na + ions into the cell is also facilitated by electrostatic interaction. As a result, the permeability of the membrane for Na + becomes 20 times greater than the permeability for K + ions.
At first, depolarization proceeds relatively slowly. When the membrane potential decreases by 10-40 mV, the rate of depolarization increases sharply and the action potential curve rises steeply. The level of membrane potential at which the rate of membrane depolarization increases sharply, due to the fact that the flow of Na + ions into the cell is greater than the flow of K + ions outward, is called critical level of depolarization.
Since the flow of Na + into the cell begins to exceed the potassium current from the cell, a gradual decrease in the resting potential occurs, leading to a reversion - a change in the sign of the membrane potential. In this case, the inner surface of the membrane becomes electropositive with respect to its outer electronegative surface. These changes in the membrane potential correspond to the ascending phase of the action potential (depolarization phase).
The membrane is characterized by increased permeability for Na + ions only for a very short time (0.2-0.5 ms). After that, the permeability of the membrane for Na + ions decreases again, and for K + - increases. As a result, the flow of Na + into the cell is sharply weakened, and the flow of K + from the cell increases.
During the action potential, a significant amount of Na + enters the cell, and K + ions leave the cell. Restoration of cellular ion balance is carried out due to the work of the sodium-potassium pump, the activity of which increases with an increase in the internal concentration of Na + ions and an increase in the external concentration of K + ions. Due to the work of the ion pump and the change in the permeability of the membrane for Na + and K +, their concentration in the intra- and extracellular space is gradually restored.
The result of these processes is the repolarization of the membrane: the inner contents of the cell again acquire a negative charge in relation to the outer surface of the membrane.
Trace negative potential is recorded during the period when Na + channels are inactivated and repolarization associated with the release of K + ions from the cell occurs more slowly than during the descending part of the action potential peak. This long-term preservation of the negativity of the outer surface of the excited area in relation to the unexcited one is called after depolarization. Trace depolarization means that during this period the outer surface of the excitable formation has a lower positive charge than at rest.
Trace positive potential corresponds to the period of increase in the resting membrane potential, i.e. membrane hyperpolarization. During a trace positive potential, the outer surface of the cell is more positively charged than at rest. The trace positive potential is often referred to as the trace hyperpolarization. It is explained by the long-term preservation of increased permeability for K + ions. As a result, a potential equal to the equilibrium potential is established on the membrane (for K + - 90 mV).
Changes in excitability in the process of development of excitation. Influencing stimuli of different strengths in different phases of the action potential, one can trace how excitability changes during excitation. On fig. 2.13" it can be seen that the period of the local response is characterized by increased excitability (the membrane potential approaches the critical level of depolarization); during the depolarization phase, the membrane loses excitability (the cell becomes refractory), which is gradually restored during repolarization.
Allocate absolute refractory period, which lasts about 1 ms in nerve cells and is characterized by their complete non-excitability. The period of absolute refractoriness occurs as a result of the almost complete inactivation (impermeability) of sodium channels and an increase in the potassium conductivity of the membrane. Even at rest, not all membrane channels are activated, 40% of them are in a state of inactivation. During depolarization, the number of inactivated channels increases and the peak of the action potential peak corresponds to the inactivation of all sodium channels.
As the membrane repolarizes, sodium channels are reactivated. it relative refractory period: an action potential can only arise under the action of stronger (above-threshold) stimuli.
AT period of negative trace potential the phase of relative refractoriness is replaced by a phase of increased (supernormal) excitability. During this period, the irritation threshold is reduced compared to the initial value, since the membrane potential is closer to the critical value than at rest (Fig. 2.14).
The phase of trace hyperpolarization, due to the residual release of potassium from the cell, on the contrary, is characterized by a decrease in

Rice. 2.13.
A - components of the excitation wave: 1 - depolarization; 2 - repolarization; MP - membrane potential; mV - microvolt; MC - critical level of depolarization: a - threshold potential duration; b - duration of the action potential; c - trace negativity; d - trace positivity; B - changes in excitability in different phases of the excitation wave; HC - the level of excitability at rest: a - an increase in excitability during the period of threshold potential; b - drop in excitability to zero during the course of the action potential (absolute refractoriness); c, - return of excitability to the initial level during trace negativity (relative refractoriness); in 2 - an increase in excitability during the period of the end of trace negativity (exaltation "or supernormality); in - the entire period of trace negativity; d - a decrease in excitability during the period of hyperpolarization (subnormality)
excitability. Since the membrane potential is greater than at rest, a stronger stimulus is required to "shift" it to the level of critical depolarization.
Thus, in the dynamics of the excitatory process, the ability of the cell to respond to stimuli changes, i.e. excitability.

Rice. 2.14.
The value of the membrane potential: E 0 - at rest; - in the exaltation phase; E 2 - in the phase of hyperpolarization. The value of the threshold potential: e 0 - at rest; e, - in the phase of exaltation; e 2 - in the phase of hyperpolarization
This is of great importance, since at the moment of the greatest excitation (peak of the action potential), the cell becomes absolutely non-excitable, which protects it from death and damage.
- See: Leontyeva N.N., Marinova K.V. Decree. op.
- There.
Lecture 2. General physiology of excitable tissues. Resting potential. action potential.
۩ The essence of the excitation process. The essence of the excitation process can be formulated as follows. All cells of the body have an electrical charge, which is created by an unequal concentration of anions and cations inside and outside the cell. Different concentrations of anions and cations inside and outside the cell is a consequence of the unequal permeability of the cell membrane for various ions and the operation of ion pumps. The process of excitation begins with the action of an irritant on an excitable cell. First, the permeability of its membrane for sodium ions increases very quickly and quickly returns to normal, then for potassium ions and also quickly, but with some delay returns to normal. As a result, ions move in and out of the cell according to the electrochemical gradient - this is the process of excitation. Excitation is possible only if the cell constantly maintains the resting potential (membrane potential) and when it is stimulated, the permeability of the cell membrane changes rapidly.
۩ resting potential. Resting potential (RP) - this is the difference in electrical potentials between the internal and external environments of the cell at rest. In this case, a negative charge is registered inside the cell. The value of PP in different cells is different. So, in the fibers of the skeletal muscle, a RI equal to 60-90 mV is recorded, in neurons - 50-80 mV, in smooth muscles - 30-70 mV, in the heart muscle - 80-90 mV. Cell organelles have their own variable membrane potentials.
The immediate reason for the existence of the resting potential is the unequal concentration of anions and cations inside and outside the cell (see tab. 1!).
Table 1. Intra- and extracellular concentrations of ions in muscle cells.
|
Intracellular concentration, mM |
Extracellular concentration, mM |
|||
|
A- (large molecular intracellular anions) |
A- (large molecular intracellular anions) |
|||
|
A small amount of |
A small amount of |
|||
|
Very little |
Basic Quantity |
|||
The uneven arrangement of ions inside and outside the cell is a consequence of the unequal permeability of the cell membrane for various ions and the operation of ion pumps that transport ions into and out of the cell contrary to the electrochemical gradient. Permeability - this is its ability to pass water, uncharged and charged particles according to the laws of diffusion and filtration. It is defined:
Channel sizes and particle sizes;
The solubility of particles in the membrane (the cell membrane is permeable to lipids soluble in it and impermeable to peptides).
Conductivity - is the ability of charged particles to pass through the cell membrane according to the electrochemical gradient.
Different permeability of various ions plays an important role in the formation of PP:
Potassium is the main ion responsible for the formation of PP, since its permeability is 100 times higher than that for sodium. With a decrease in the concentration of potassium in the cell, PP decreases, and with an increase, it increases. He can go in and out of the cell. At rest, the number of incoming potassium ions and outgoing potassium ions is balanced and the so-called potassium equilibrium potential is established, which is calculated according to the Nernst equation. Its mechanism is as follows: since the electrical and concentration gradients oppose each other, potassium tends to go outside along the concentration gradient, and a negative charge inside the cell and a positive charge outside the cell prevents this. Then the number of incoming ions becomes equal to the number of outgoing ions.
Sodium enters the cell. Its permeability is small compared to the permeability of potassium, so its contribution to the formation of PP is small.
Chlorine enters the cell in an insignificant amount, since the permeability of the membrane for it is small, and it is balanced by the amount of sodium ions (opposite charges attract). Therefore, its contribution to the formation of PP is small.
Organic anions (glutamate, aspartate, organic phosphates, sulfates) cannot leave the cell at all, since they are large. Therefore, due to them, a negative charge is formed inside the cell.
The role of calcium ions in the formation of PP is that they interact with the external negative charges of the cell membrane and the negative carboxyl groups of the interstitium, neutralizing them, which leads to the stabilization of PP.
In addition to the above ions, the surface charges of the membrane (mostly negative) also play an important role in the formation of PP. They are formed by glycoproteins, glycolipids, and phospholipids: fixed external negative charges, neutralizing the positive charges of the outer surface of the membrane, reduce the RI, and fixed internal negative charges of the membrane, on the contrary, increase the RI, summing up with anions inside the cell. In this way, resting potential is the algebraic sum of all positive and negative charges of ions outside and inside the cell and the surface charges of the cell membrane.
The role of ion pumps in the formation of PP. ion pump is a protein molecule that ensures the transfer of an ion with a direct expenditure of energy, contrary to the electrical and concentration gradients. As a result of conjugated transport of sodium and potassium, a constant difference in the concentrations of these ions inside and outside the cell is maintained. One ATP molecule provides one cycle of the Na / K-pump - the transfer of three sodium ions outside the cell and two potassium ions inside the cell. Thus, the PP increases. The normal value of the resting potential is a necessary condition for the formation of an action potential, that is, for the formation of the excitation process.
۩action potential. action potential - This is an electrophysiological process, which is expressed in a rapid fluctuation of the membrane potential due to a change in the permeability of the membrane and the diffusion of ions into and out of the cell. Role of PD is to ensure the transmission of signals between nerve cells, nerve centers and working organs, in the muscles PD provides the process of electromechanical pairing. PD obeys the all-or-nothing law. If the force of irritation is small, then a local potential arises, which does not propagate.
The action potential consists of three phases: depolarization, that is, the disappearance of PP; inversions - changes in the sign of the cell charge to the opposite; repolarization - restoration of the original MP.
Action Potential Mechanism.
Depolarization phase . Under the action of an irritant on a cell, the initial partial depolarization of the cell membrane occurs without changing its permeability to ions. When depolarization reaches about 50% of the threshold value, the permeability of the membrane for Na + increases, and at the first moment relatively slowly. During this period, the driving force that ensures the movement of Na + into the cell is the concentration and electrical gradients. Recall that the cell inside is negatively charged (opposite charges attract), and the concentration of Na + outside the cell is 12 times greater than inside the cell. The condition for further entry of Na + into the cell is an increase in the permeability of the cell membrane, which is determined by the state of the gate mechanism of sodium channels. The gate mechanism of sodium channels is located on the outer and inner side of the cell membrane, the gate mechanism of potassium channels is located only on the inner side of the membrane. The sodium channels have activation m-gates, which are located on the outer side of the cell membrane, and inactivation h-gates, located on the inner side of the membrane. At rest, the activation m-gates are closed, and the inactivation h-gates are open. The potassium activation gate is closed, but there is no inactivation potassium gate. When the cell depolarization reaches a critical value, which is usually 50 mV, the permeability of the membrane for Na + increases sharply, since a large number of voltage-dependent m-gates of sodium channels open and sodium ions rush into the cell in an avalanche. The developing depolarization of the cell membrane causes an additional increase in its permeability and, accordingly, sodium conductivity: more and more activation m-gates open. As a result, the PP disappears, that is, it becomes equal to zero. The depolarization phase ends here. Its duration is approximately 0.2-0.5 ms.
Phase inversion . The process of membrane recharging is the second phase of PD, the inversion phase. The inversion phase is divided into ascending and descending components. ascending part . After the disappearance of PP, the entry of sodium ions into the cell continues, since the sodium activation m-gate is still open. As a result, the charge inside the cell becomes positive, and outside - negative. For a fraction of a millisecond, sodium ions still continue to enter the cell. Thus, the entire ascending part of the AP peak is provided mainly by the entry of Na + into the cell. Downward component of the inversion phase . Approximately 0.2-0.5 ms after the onset of depolarization, the growth of AP stops as a result of the closing of the sodium inactivation h-gate and the opening of the potassium activation gate. Since potassium is located mainly inside the cell, it, according to the concentration gradient, begins to quickly leave it, as a result of which the number of positively charged ions in the cell decreases. The charge of the cell starts to decrease again. During the descending component of the inversion phase, the release of potassium ions from the cell is also facilitated by an electrical gradient. K+ is pushed out by the positive charge from the cell and is attracted by the negative charge outside the cell. This continues until the complete disappearance of the positive charge inside the cell. Potassium leaves the cell not only through controlled channels, but also through uncontrolled channels - leakage channels. The AP amplitude is the sum of the PP value and the inversion phase value, which is 10-50 mV in different cells.
Repolarization phase . As long as the activation potassium channels are open, K+ continues to flow out of the cell, according to the chemical gradient. The charge inside the cell becomes negative, and outside - positive, therefore, the electrical gradient sharply inhibits the release of potassium ions from the cell. But since the strength of the chemical gradient is greater than the strength of the electrical gradient, potassium ions continue to leave the cell very slowly. Then the activation potassium gate closes, leaving only the release of potassium ions through the leakage channels, that is, along the concentration gradient through uncontrolled channels.
Thus, PD is caused by a cyclic process of sodium ions entering the cell and subsequent potassium exit from it. The role of Ca 2+ in the occurrence of AP in nerve cells is insignificant. However, Ca 2+ plays a very important role in the occurrence of AP of the heart muscle, in the transmission of impulses from one neuron to another, from a nerve fiber to a muscle fiber, and in ensuring muscle contraction.
AP is followed by trace phenomena (characteristic of neurons), first trace hyperpolarization and then trace depolarization. trace hyperpolarization cell membrane is usually a consequence of the still remaining increased permeability of the membrane for potassium ions. trace depolarization associated with a short-term increase in membrane permeability for Na + and its entry into the cell according to chemical and electrical gradients.
In addition, there are: a) the so-called phase absolute refractoriness, or complete non-excitability of the cell. It falls on the peak of AP and will last 1-2 ms; and b) relative refractoriness phase- a period of partial recovery of the cell, when a strong irritation can cause a new excitation. Relative refractoriness corresponds to the final part of the repolarization phase and trace hyperpolarization of the cell membrane. In neurons, after hyperpolarization, partial depolarization of the cell membrane is possible. During this period, the next action potential can be caused by weaker irritation, since the MP is somewhat less than usual. This period is called exaltation phase(period of increased excitability).
The rate of phase changes in cell excitability determines its lability. Lability, or functional mobility, is the rate of one excitation cycle. A measure of the lability of an excitable formation is the maximum number of AP that it can reproduce in 1 second. Typically, the excitation lasts less than 1 ms and is like an explosion. Such an "explosion" proceeds powerfully, but quickly ends.
Muscles, differences Document
... . Excitability fabrics and its measure. Laws of irritation excitable fabrics: strength, time actions irritant... potential rest(MPP); 2) membrane potential actions(IPD); 3) potential basal metabolic rate (metabolic potential). Potential ...
It is restored due to diffusion from the cell of positively charged potassium ions, the concentration of which in the environment is also much lower than the intracellular one.
Encyclopedic YouTube
1 / 5
Electrotonic potentials and action potentials
Action potential of cardiomyocytes
Action potential of the working myocardium
Action potential in pacemaker cells
Membrane Potentials - Part 1
Subtitles
In the previous video, we talked about how the cell uses the sodium-potassium pump and ATP to maintain the potential difference between the inside of the cell and the outside. In general, the external environment is more positively charged than the internal one. We have a potential difference of -70 millivolts between the internal and external environment. This is a minus value because the environment is more positively charged. If a less positive value is subtracted from a more positive value, then you get a negative value of the order of -70. This statement is the basis for understanding how neurons transmit signals in the body. And to better explain this, I want to give you one more concept in addition. After that, you will understand what is the principle of signal transmission by a neuron in real life. And what's more, you will understand why they need these myelin sheaths and nodes of Ranvier, and what all these dendrites are for. I hope you get a complete picture. So there are two ways in which the potential can move. Two ways of signal transmission. Let's call it that.. I don't know which word fits better. So, the first way is electrotonic. It sounds very complicated, but you will see that the idea behind it is very simple. Let me write down the term first. Electrotonic... electrotonic potential. And the second thing I'm going to focus on is the action potential. Both of them have positive and negative sides in terms of signal transmission capability. We will talk about them within the framework of ideas about the cell and the cell membrane. Let's now deal with all this in detail. So, I will draw the cell membrane. Let's agree that this is a nerve cell, or a neuron, because we are discussing precisely nerve cells. We know that the more positive charge is on the outside. We also know that there is a lot of sodium on the outside, and there is more sodium on the outside than on the inside. There may be very little of it here. And we know that the inside contains a lot more potassium than the outside, however we also know that the outside has a greater positive charge than the inside because our sodium-potassium pump will pump out three sodium ions for every two potassium ions it pumps. inside. And in the last video, I told you that there are structures called sodium gates. Gate for sodium ions. Here are all the ions. They have a charge. And now let's say that there are some circumstances, some incentives - let me point them out. I'll go a little lower. This is where our sodium ion gate is located. They are in the closed position, but let's say something is causing them to open. We'll talk maybe in this video, or in this and the next video about the various things that can cause them to open. A certain type of stimulus triggers their opening. In fact, there are a number of different incentives. In general, let's say they opened. What happens after their discovery? We opened them, or rather, a certain stimulus did it - what will happen next? We have more positive charge on the outside than on the inside, so positively charged objects will want to get in. And it's a sodium gate, so only sodium can get through it. This is a kind of curved protein structure through which only sodium ions can penetrate. And on top of everything else, we have a lot more sodium on the outside than on the inside. Therefore, the diffusion gradient will encourage sodium to pass through it. And as a result of the fact that sodium ions are positively charged, and the environment is also more positively charged, they will tend to leave this positively charged environment. So, if you open this gate, you will have a lot of sodium ions ready to come in. I'll draw more of them here. And now, what should happen if we move further along the membrane? Let's enlarge the image. So let's imagine that our membrane. And this is our open gate, for some reason they opened, and a large amount of sodium ions penetrate inside. And it all becomes more positively charged. Let's pretend we have a voltmeter here. We measure the potential difference between the inner surface of the membrane and the outer surface. Let me draw a little diagram. I'll draw a circuit right here on my voltmeter. And that would be the potential difference - or we can call it the membrane voltage, or the voltage difference between the two membrane surfaces - and let's label this axis as time. Imagine that we have not yet opened this gate. That is, it is a state of rest. Our sodium-potassium pumps continue to work. Particles leak back and forth, but the ratio is held at minus 70 millivolts. So, here we have minus 70 millivolts. And now, as soon as these gates, which are located in some other parts of the cell, open, what happens? Assume, however, that only these gates open. So here suddenly a more positive charge is formed. So positive charges are already there - so other positive charges due to sodium or potassium ions will tend to be as far away from this point as possible, because there is no flow of positive ions in this zone. That is, it is less positively charged compared to this area. We have some potassium ions here, and maybe some sodium ions. All of them will strive to be as far as possible from the place where the gate opened. The charge will tend to spread as far as possible. And as soon as this happens, as soon as we open this gate, we have a shift of positive charge in this direction. This happened at minus 70 millivolts. So, some positive charge goes away. All this happens very quickly. Almost immediately after that, the charge becomes less negative, or more positive. The potential difference between this and this areas becomes smaller. This corresponds to a given point on the graph. And now, if we take this point, if we do the same - if we measure the voltage at this point right here, then maybe it will be minus 70 millivolts, and after a while, the positive charge will begin to take its toll and the charge here will become more positive, while the effect will decrease. Because these positive charges will spread in all directions. As a result, the effect seems to be rarefied. That is, it becomes less pronounced. And the charge here will become less positive. So, the electrotonic potential occurs at one point in the cell, when the gate opens and the charge begins to spread inward, and begins to affect the potential in other parts of the cell. However, the positive thing is that it is very fast. As this happens... As it enters the cell, it becomes more and more positive, but the farther it goes, the effect is more and more dissipated... more and more dissipated as the distance increases. And if speed is important to you, then you need exactly this electrotonic potential. As soon as this happens, all other parts of the cell are affected, but if you want this change in potential to propagate over longer distances - for example, let's decide that if we went all the way to this point of the neuron and now want to measure it, then it won't have any effect. Perhaps a little later, but this potential will have no effect, since the charge is dissipated as the charge increases in the entire cell. So, this influence is away from the original localization where the opening of the gate took place. It will be significantly less. Distance doesn't really help the action. And now let's try to decide what will happen to the action potential. From the name it is clear that in this case there will be more action. So let's start with the same situation. We have sodium gates that open in response to certain stimuli. And now I will draw two membranes. Here is the outside. And, accordingly, this is the inner side. Let me draw a picture - perhaps we've come across this before - but now we'll break it down in more detail. Let's say this is an axon, and let me draw another sodium gate here. They are very close. And they must be transformable. So they transform, and here I have another sodium gate. I will draw them too. But I don't think it's necessary to draw them in large numbers. You need to draw one cluster so that you understand what is happening. Let me draw another potassium gate. Good. All gates are drawn. Let's agree that they are all initially closed. So, they are all in the closed position. And now that sodium gate is being stimulated. And they open up. Yes, let's say these gates open. They are stimulated by something and open up as a result. Let's decide that specifically these sodium gates open under the influence of a certain stimulus that has a certain voltage. Let the gate open when it reaches minus 55 millivolts. I'll write it down. And when they are at rest, the potential difference between the inside of the cell and its outside is minus 70 millivolts, and therefore they don't open. They remain closed, however, if under certain circumstances this charge becomes positive enough to provide a value of minus 55 millivolts, these gates open. Let's write down a few rules that govern what happens to this gate. They close - and these are all just ballpark numbers, the main goal is for you to get the basic idea. Let them close at plus 35 millivolts. And our potassium gates open at plus 40 millivolts, just so you get the basic idea. And the potassium gate closes at minus 80 millivolts. So be it. And what happens as a result? Let's assume that, for some reason, the voltage here becomes minus 55 millivolts. I will now draw a diagram, by analogy with how I did it before. So, I need some space to draw my diagram. So the pattern is the same. This is the membrane voltage. Good. But this time. And let's pretend we're measuring voltage - let's say it's membrane voltage near the sodium gate right here. So, we measure the voltage on both sides of the membrane. And if there is no stimulation, then we get a value of the order of minus 70 millivolts - and suddenly some kind of stimulus, for some reason, makes this area more positively charged. Perhaps this is a certain type of electrotonic effect that makes this area more positively charged. Perhaps there are certain positive charges here. And, as a result, this area becomes more positively charged. Then the sodium-potassium pumps pump the ions out, causing the minus 55 millivolt threshold to be reached, resulting in nothing happening, right? However, when there is another electrotonic stimulus, or maybe several, a large amount of positive charges are concentrated here, resulting in a value of minus 55 millivolts. Remember that as soon as there is a positive charge, everything becomes less negatively charged. And the potential difference is getting smaller. And when we reach minus 55 millivolts, the gate opens. Here they were closed before. They were closed at minus 70 millivolts. So let me write it down here. At this point, our sodium gate opens. And now what happens when our sodium gate opens? When they open - we've seen this before - all the positively charged sodium ions go over here, both in the direction of the electrical gradient and the diffusion gradient, and get inside the cell. There's so much sodium out here, so much positive charge in here, they're just trying to get in. And once they reach that threshold, even though it can only happen at minus 55 or maybe minus 50 millivolts, the result is that the gate opens and all of our positive charge enters the cell. And the potential difference becomes much more positive. Okay, now move on. They keep coming in and the charge gets more and more positive, and as it gets more positive, the gate closes at plus 35 millivolts. Now I'll free up more space to continue. So let's pretend we're here - let's say we have 35 millivolts plus here. The gate closes, and at the same time, all this that I just removed - I set it to plus 40 millivolts, no, plus 35, just to support my idea. So let's imagine that at plus 35 millivolts our sodium gate opens. What happens as a result? All of a sudden we're at plus 35, or maybe plus 40 millivolts like this - let's say plus 40, I guess you get the idea anyway, so let's say plus 40. So, at plus 40, these gates close. No more positively charged ions come in, but now we have a more positive charge on the inside, at least locally at this point on the membrane, than on the outside. And those gates are opening. So, as a result, our potassium gate opens. K-plus ionic gates open. And what happens now that they are open? We have all these sodium ions here. We have already seen on the example of the sodium-potassium pump that potassium - all these potassium ions are located here. We have seen with the sodium-potassium pump that it increases the sodium concentration on the outside, while the potassium concentration is higher inside the cell. And now, in addition to this positive charge of plus 40 millivolts, we also have a more positive charge on the inner surface of the membrane. So they open up. And these particles want to leave because there are fewer potassium ions outside. And they want to follow their concentration gradient. This side also has a large positive charge. Approximately plus 40 millivolts. The ions want to be released. They start to leave the cage. So, positive charges begin to leave the cell from the inside out. And we become less positively charged again. Let me write down what is happening here. So at this point our sodium gate closes and our potassium gate opens. Some closed, others opened. After that, the positive charges begin to exit the cell again, and perhaps I exaggerated a little, because they will close, probably when we already get minus 80 millivolts. Let's say the potassium gate closes at -80. And then our sodium-potassium pump can bring us back to our minus 70 millivolts. So, this is what happens in this area of the cell, in the immediate vicinity of the first sodium gate. However, what happens in general? As a very positive charge builds up here, we're getting close to 40 millivolts right here. We have reached 40 millivolts in this area of the cell. As you have practically observed, over a short distance of the electrotonic potential, this area becomes more positively charged. It becomes more positively charged. These positive charges begin to spread to where the charge is less positive. That is, it becomes more positive. It was at minus 70 millivolts, but it's getting more positive. Gradually it increases to minus 65, minus 60, minus 55 and then “Bam!”. Again there is stimulation. And those gates are opening again. These gates are opening again. Sodium re-enters. And if you want to follow this gate, the difference in potentials of what goes through it, all this happens as soon as sodium starts flowing through this first gate, through the second gate - it potentiates here, because the second gate is located a little later in time - because all this flow passes a little to the left of it, its potential increases. He potentiates, and the same thing happens to him again. When sodium comes in here, the environment becomes positively charged, and this causes the cell, the voltage around, the charge to become a little more positive, and this triggers the opening of the next sodium gate and then the same thing happens, the same cycle repeats. The potassium gate then opens to allow the negative charge to return, however by the time this happens the environment is already positively charged enough to trigger another sodium gate. So, one after the other, these sodium gates open and close, so the information is transmitted, this change in potential is transmitted. So what's going on here? So, it happens more slowly and with the attraction of energy. It happened - electrotonic potential - very quickly. And this process is slow. The action potential is slower. I don't mean that it is completely slow. It's slower because it has to involve all those gate openings and closings, and it also requires energy. I'll write it down. And you also have to constantly provide energy to the potential in our cell, and use your sodium-potassium pumps, which are in an active state. But it's good. The positive side is that it covers the distance well. Let's write this down too. We have observed with the example of the electrotonic potential that as we move farther and farther away from where the stimulation occurred, the change in potential becomes more and more dispersed. It fades exponentially. It gets more and more scattered as we go further, which is not good for long distances. This can go on indefinitely, because each time a new gate is stimulated, and that gate continues to provide a flow of ions, incoming ions, as well as those that make the environment a little more negatively charged. This is followed by the opening of the next gate. And this allows you to travel long distances very efficiently. And now we really have a foundation for understanding what's going on in a neuron, and I'll continue this topic in the next video, where I'll show you how electrotonic potentials and action potentials can combine to provide a signal through the neuron.
Action potential phases
- prespike- the process of slow depolarization of the membrane to a critical level of depolarization (local excitation, local response).
- Peak potential, or spike, consisting of an ascending part (membrane depolarization) and a descending part (membrane repolarization).
- Negative trace potential- from the critical level of depolarization to the initial level of membrane polarization (trace depolarization).
- Positive trace potential- an increase in the membrane potential and its gradual return to its original value (trace hyperpolarization).
General provisions
The polarization of the membrane of a living cell is due to the difference in the ionic composition of its inner and outer sides. When the cell is in a calm (unexcited) state, ions on opposite sides of the membrane create a relatively stable potential difference, called the resting potential. If an electrode is inserted into a living cell and the resting membrane potential is measured, it will have a negative value (of the order of -70 - -90 mV). This is explained by the fact that the total charge on the inner side of the membrane is significantly less than on the outer one, although both sides contain both cations and anions. Outside - an order of magnitude more sodium, calcium and chlorine ions, inside - potassium ions and negatively charged protein molecules, amino acids, organic acids, phosphates, sulfates. It must be understood that we are talking about the charge of the membrane surface - in general, the environment both inside and outside the cell is neutrally charged.
The membrane potential can change under the influence of various stimuli. An artificial stimulus can be an electrical current supplied to the outer or inner side of the membrane through the electrode. Under natural conditions, the stimulus is often a chemical signal from neighboring cells, coming through the synapse or by diffuse transmission through the intercellular medium. The shift of the membrane potential can occur in the negative ( hyperpolarization) or positive ( depolarization) side.
In nervous tissue, an action potential, as a rule, occurs during depolarization - if the depolarization of the neuron membrane reaches or exceeds a certain threshold level, the cell is excited, and an electrical signal wave propagates from its body to axons and dendrites. (In real conditions, postsynaptic potentials usually arise on the neuron body, which are very different from the action potential in nature - for example, they do not obey the “all or nothing” principle. These potentials are converted into an action potential at a special section of the membrane - the axon hillock, so that action potential does not extend to dendrites).
This is due to the fact that there are ion channels on the cell membrane - protein molecules that form pores in the membrane through which ions can pass from the inside of the membrane to the outside and vice versa. Most channels are ion-specific - the sodium channel passes practically only sodium ions and does not pass others (this phenomenon is called selectivity). The cell membrane of excitable tissues (nerve and muscle) contains a large amount of potential-dependent ion channels capable of rapidly responding to shifts in membrane potential. Membrane depolarization primarily causes voltage-gated sodium channels to open. When enough sodium channels open at the same time, positively charged sodium ions rush through them to the inside of the membrane. The driving force in this case is provided by the concentration gradient (there are many more positively charged sodium ions on the outside of the membrane than inside the cell) and the negative charge on the inside of the membrane (see Fig. 2). The flow of sodium ions causes an even larger and very rapid change in the membrane potential, which is called action potential(in the special literature it is designated PD).
According to all-or-nothing law the cell membrane of an excitable tissue either does not respond to the stimulus at all, or responds with the maximum possible force for it at the moment. That is, if the stimulus is too weak and the threshold is not reached, the action potential does not arise at all; at the same time, the threshold stimulus will elicit an action potential of the same amplitude as the stimulus above the threshold. This does not mean that the amplitude of the action potential is always the same - the same section of the membrane, being in different states, can generate action potentials of different amplitudes.
After excitation, the neuron for some time finds itself in a state of absolute refractoriness, when no signals can excite it again, then enters a phase of relative refractoriness, when exceptionally strong signals can excite it (in this case, the AP amplitude will be lower than usual). The refractory period occurs due to the inactivation of the fast sodium current, i.e. the inactivation of the sodium channels (see below).
Action potential propagation
On unmyelinated fibers
In the course of PD, the channels pass from state to state: Na + channels have three basic states - closed, open and inactivated (in reality, the matter is more complicated, but these three are enough for a description), K + channels have two - closed and open.
The behavior of the channels involved in the formation of TP is described in terms of conductance and calculated in terms of transfer (transfer) coefficients.
The transfer coefficients were derived by Hodgkin and Huxley.
Conductivity for potassium G K per unit area
| d n / d t = α n (1 − n) − β n n (\displaystyle dn/dt=\alpha _(n)(1-n)-\beta _(n)n), |
| where: |
| α n (\displaystyle \alpha _(n))- transfer coefficient from closed to open state for K+ channels ; |
| β n (\displaystyle \beta _(n))- transfer coefficient from open to closed state for K+ channels ; |
| n (\displaystyle n)- fraction of K+ channels in the open state; |
| (1 − n) (\displaystyle (1-n))- fraction of K+ channels in the closed state |
is more difficult to calculate, because, as already mentioned, for voltage-dependent Na+ channels, in addition to the closed/open states, the transition between which is described by the parameter m (\displaystyle m), there is also an inactivated/non-inactivated state, the transition between which is described through the parameter h (\displaystyle h)
| d m / d t = α m (1 − m) − β m m (\displaystyle dm/dt=\alpha _(m)(1-m)-\beta _(m)m), | d h / d t = α h (1 − h) − β h h (\displaystyle dh/dt=\alpha _(h)(1-h)-\beta _(h)h), |
| where: | where: |
| α m (\displaystyle \alpha _(m))- transfer coefficient from closed to open state for Na+ channels ; | α h (\displaystyle \alpha _(h))- transfer coefficient from inactivated to non-inactivated state for Na+ channels ; |
| β m (\displaystyle \beta _(m))- transfer coefficient from open to closed state for Na+ channels ; | β h (\displaystyle \beta _(h))- transfer coefficient from non-inactivated to inactivated state for Na+ channels ; |
| m (\displaystyle m)- fraction of Na+ channels in the open state; | h (\displaystyle h)- fraction of Na+ channels in non-inactivated state; |
| (1 − m) (\displaystyle (1-m))- fraction of Na+ channels in the closed state | (1 − h) (\displaystyle (1-h))- fraction of Na+ channels in the inactivated state. |
The combination of organic matter and metals in living beings, the use of a living cell as a conductor of electric current cannot but surprise. The more details about my body I learn, the more often there is surprise from its simple actions. Bending the arm can be perceived not as a habitual, almost unnoticed process, but as a result of the coordinated work of muscles and tendons that received a command from the brain, transmitted through nerve fibers with the help of electricity. Looking at the band of deep iridescent blue that appears on a piece of polished labradorite that I turn in my hands, I perceive the electrical waves emitted by it with the receptors of my eyes. In receptors, this information is encoded in the form of a series of electrical impulses. It is transmitted to the brain, processed by it, and the result of all this is the perception of a mysteriously beautiful stone, the emotions that arise during this. Matrix, in short :)
Along with admiration, there is also a desire to find out how it all happens, a desire to delve deeper into the details of each process.
An action potential is that electrical impulse that delivers information from receptors to the brain, and with the help of which the brain controls the body. Its existence requires specially arranged cells, neurons, the membranes of which are plastered on both sides with positively and negatively charged ions, which are just waiting for a signal in order to carry this electrical impulse along the entire length of the cell. The main place of existence of the action potential is the axon of the neuron. The dendrites of some types of neurons are also capable of conducting electrical impulses.

It is enough to look at the picture on the left to see the main difference between the resting potential and the action potential: when the membrane is at rest, there is a positive electric charge on its entire length on the outside, and a negative one on the inside.

And the action potential passing through the neuron from the cell body to the end of the axon is a change in the external and internal charges of the cell membrane in a small area of the neuron: for a very short period of time, the negative and positive charges on opposite sides of the membrane change places.
Questions arise: what triggers an action potential? What is its physical nature, what exactly happens in the cell during its existence? How does it spread? Why does it only go in one direction? How does a cell restore its initial resting potential after a wave of excitation has passed through it?
So the first question is: How is an action potential generated? Since the transmission of signals from source neurons to the recipient neuron, or postsynaptic neuron (postsynaptic neuron), occurs in synapses, the signal that triggers the action potential should be looked for there. Through the synapse, signaling substances come to the neuron, but their arrival does not necessarily mean the triggering of the excitation of the neuron. If the neuron responded to very small amounts of signaling substances, it would be as inconvenient as having a super-sensitive switch that works if you accidentally sneeze on it.

It takes a strong enough exposure to certain signaling substances to trigger an action potential. In real conditions, postsynaptic potentials usually arise on the body of a neuron - temporary changes in the charges of the membrane caused by signal substances that have come through the synapse.
Post-synaptic potentials are very different from action potentials in nature. If the action potential has one strictly defined value, then the value of postsynaptic potentials depends only on the type and amount of chemical substances that caused them, transmitted through the synapses. While the action potentials generated by different neurons are approximately the same, the postsynaptic potentials that occur at different input synapses on the same neuron differ greatly in magnitude and duration. In one synapse of a neuron, an incoming nerve impulse can cause a change in the membrane potential by 0.1 mV, and in another, by 20 mV. In addition, there are both action potential-causing and inhibitory signals.
The cell body serves as a kind of calculator, the simplest analog computer that sums up all the signals entering it in the form of emerging postsynaptic potentials. Analog computers are very simple. For example, you can build a water analog computer. If three kilograms of water are poured into a jar from one pipe, and five kilograms of water from another into the same jar, then after weighing the jar and finding out that the scales show a weight equal to eight kilograms, we conclude that 3 + 5 = 8. At the dawn of the development of computer technology, both water and other analog computers were used, but the most advanced was, of course, a machine that sums up electric currents exactly like neurons. The operators of such computers were engaged in calculations, sticking plugs from one socket to another, merging different currents together.
So, in a neuron, the same summation of electric currents occurs. In the process of this summation, one can go over excitation threshold (excitation threshold): a certain value of the membrane potential will be reached, which triggers the action potential. Pay attention to the fact that to reach the threshold of excitation, the membrane potential must increase. If the resting membrane potential of a neuron is -70 mV, then its excitation threshold can be -55 mV. A decrease in the membrane potential, for example, to -90 mV leads to the fact that a stronger effect is required to overcome the excitation threshold, because it remains the same: -55 mV.

In a postsynaptic neuron that receives signals from two excitatory and one inhibitory neurons, there may be various combinations of signals, and not every one of them leads to the occurrence of an action potential.
In the first diagram of the picture above, a situation is depicted when a weak signal comes from only one excitatory neuron, which is not enough to overcome the threshold of excitation. This signal is repeated, but by the time a second, equally weak signal arrives from the same axon, the excitation of the membrane has already reached the resting potential, and no action potential arises.
The second diagram shows the same weak signal, but coming at a higher frequency. It is because of the increase in the frequency of the signal that the membrane does not have time to return to a state of rest in the interval between two signals. In this case, the sum of the residual charge of the membrane after the first signal and the second signal coming after it makes it possible to overcome the excitation threshold of the neuron. In this case, at the base of the axon, which is called axon hillock (axon hillock [‘aksɒn ‘hɪlək]), an action potential occurs.
In the third scheme, weak signals come from both excitatory neurons, and their sum is enough to trigger an action potential.
The fourth scheme shows the effect of excitatory and inhibitory signals on the postsynaptic neuron. When only an excitatory signal is received, the membrane potential slightly increases, and when only an inhibitory signal is received, it decreases. If these two signals come at the same time, then they just destroy each other, leaving the membrane potential in a state close to the resting potential. This is reminiscent of pressing on the same door from different sides: separately, each impact could slightly open the door or, vice versa, close it, but existing simultaneously, they only lead to a slight shift in the direction of action of a larger force.
The nature of the system is such that even small postsynaptic potentials, summed up, can produce a large effect. The neuron “adds up” all the excitatory postsynaptic potentials that have entered it, “subtracts” the incoming inhibitory postsynaptic potentials, and as soon as the excitation threshold is crossed, an action potential arises.
To understand what happens in the cell during triggering and passing an action potential, it is necessary to list each participating player and his action, as was done when describing the occurrence of the resting membrane potential in . So, actors in creating an action potential:

1 . , which were closed when the resting membrane potential existed, opened as soon as the potential reached the threshold of excitation. Since there is a negative charge on the membrane inside the cell, at this moment the
2 . positively charged sodium ions (Na+). It was they who created a positive charge on the outer surface of the neuron membrane, and it is at this moment that they transfer it inside, leaving a negative charge on the outer membrane, which is created by negatively charged chloride ions located outside the cell.
3 . thus chloride ions (Cl -) create a negative charge on the outer surface of the membrane.

The movement of sodium ions into the cell leads to a sharp change in the charges inside and outside the neuron. The place of the negative internal charge of the membrane that exists during the rest of the neuron is occupied by the positive charge of sodium ions. And outside the cell membrane, after Na + leaves from there, Cl - can finally manifest itself, whose negative charge is no longer compensated by positively charged ions. Thus, the potential of the cell membrane suddenly increases from -70 to +40 mV. It is this increase, called depolarization (depolarization) is depicted by the sharply rising part of the peak in this picture. This increase in membrane potential is a signal to close voltage-gated sodium channels, and also opens voltage-gated potassium channels, which we have not talked about before.
4 . voltage-gated potassium channels . They exist in the neuron in addition to the potassium flow channels, which, as you probably remember from the last chapter on potential, do not have gates and provide a constant outflow of potassium from the cell, playing an essential role in the formation of the resting potential. So it turns out that there are two types of potassium channels, and now we will look at the operation of a voltage-gated potassium channel. Opening after the action potential has reached its peak, voltage-gated potassium channels dramatically increase the outflow of potassium ions from the cell. The positive charge that exists at this moment inside the neuron, formed due to the influx of Na +, actively pushes positively charged potassium ions (K +) out of the cell through both types of channels.
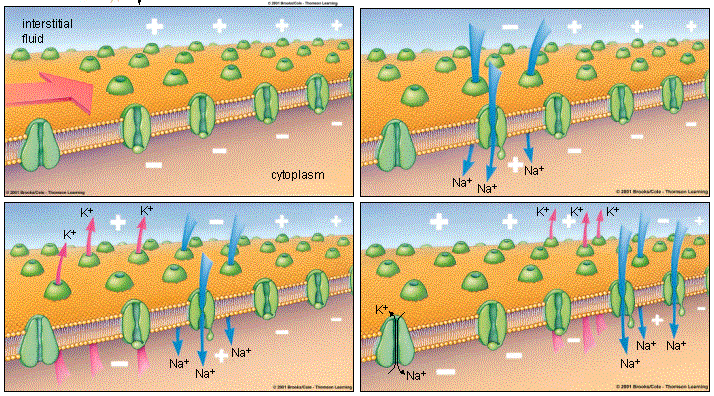
So, once again, let's trace the chain of events up to this point: the potential reaches the threshold of excitation, and this electrical signal leads to the fact that voltage-dependent sodium channels open, Na + ions rush inward, attracted by negatively charged proteins, depolarization occurs, that is, a charge difference outside and inside dropped sharply, and even more - the absence of positively charged sodium ions led to the appearance of a negative charge where there had just been positive - there was a change of poles. The action of the electric impulse ended, the sodium channels closed again, and the Na + ions crowded inside pushed out the similarly charged K + ions through the opened voltage-dependent potassium channels and through the potassium flow channels.
The charge of the inner surface of the cell membrane begins to rapidly decrease; in the diagram, this corresponds to the downward part of the peak. This decrease in charge is called repolarization (repolarization), that is, the difference in charges outside and inside the neuron is restored again, as it was initially at rest. But it was the charge difference that was restored, and not the ionic composition that created it. Despite the fact that at the end of repolarization, a positive charge is formed outside the membrane, equal to the positive charge of the membrane at rest, the membrane is still far from reaching the resting state. After all, this positive charge was brought there by K + ions that left the cell, and at rest potential it is supported by Na + ions, which are currently locked inside the cell, in the very place where potassium ions were previously at rest.
At the end of repolarization, there may be a decrease in membrane potential even below its value for a resting neuron, or hyperpolarization (hyperpolarization [ˌhaɪpəpələraɪ'zeɪʃn]). After a short period of time, it is compensated by an additional release of potassium ions from the cell, and the former, stable state of rest sets in.

When the membrane potential becomes equal to the resting potential (-70 mV), voltage-gated potassium channels close. Through the flowing potassium channels, K + no longer flows out so rapidly, because its concentration inside the cell has fallen, and there are many potassium ions outside. At the same time, there are many sodium ions inside, which must be removed from the cell in order for the action potential to pass through it again.
The sodium-potassium pump is responsible for restoring the initial position “sodium outside, potassium inside”, which removes three Na + from the cell and simultaneously brings two K + inside. His work was described in the previous chapter about the potential , and it is shown schematically in the fourth part of this picture. The combined action of the sodium-potassium pump and the potassium flow channels leads to the fact that a large concentration of Na + appears again on the outer side of the membrane, and the concentration of K + inside the neuron is higher than the outer one.
If only potassium flow channels were working, then potassium ions would only leave the cell, and it would be impossible to maintain a greater concentration of K + inside than outside. If only voltage-dependent sodium channels were active, then sodium ions could not leave the cell at all after they entered it, creating an action potential. But the sodium-potassium pump (well, that is, the sodium-potassium channel) solves both of these problems: it brings Na + out, restoring its high concentration from the outside. And it drives inside the K + that passed through the flow channels, preventing them from being distributed evenly on both sides of the membrane, thus losing the partial pressure necessary for the formation of the resting membrane potential.
A thousandth of a second after the repolarization of the neuronal membrane has reached a minimum, the active work of the membrane channels restores the resting potential. The neuron is ready to conduct the next action potential.
5 . positively charged calcium ions (Ca 2+) play an important role in the formation of the action potential only in some nerve cells, therefore, in this chapter, which contains information common to various neurons, it will not be described.

This completes the general description of the movement of ions across the cell membrane, which turns the resting potential of the neuron into an action potential, is completed. You can take a break and digest it along with something tasty, at the same time nourishing the brain, compensating for the calories that it spent on processing all this information.
Action potential propagation along the entire length of the axon, it is provided by the fact that the electric current that arose during its existence in one section of the neuron membrane excites neighboring sections, opening sodium voltage-dependent channels and triggering an action potential already there. More precisely, the electric current triggers an action potential only on one side of the place where the previous potential existed. Because, on the other hand, sodium channels are inactivated after a recent discovery, and without their participation, the occurrence of an action potential is impossible. If it were not for this time gap of non-functionality between two sodium channel openings, the action potential could not propagate in only one direction. The resulting chain reaction of sodium channel opening ensures the transmission of the nerve impulse from its origin to the axon endings suitable for other neurons (or a muscle cell).

Schematically, this chain reaction can be depicted as follows:
Time 1: The electric current that arose as a result of the opening of voltage-dependent sodium channels in section 1 excites neighboring sections of the membrane.
Point in time 2: As a result of this excitation, voltage-dependent sodium channels open in site 2 and an action potential arises, which, in turn, excites neighboring sections of the membrane. In site 1, voltage-dependent potassium channels opened, restoring the membrane potential after the passage of the action potential.
Point in time 3: The electrical current resulting from the opening of voltage-gated sodium channels in site 2 caused the opening of the same channels in site 3, and the action potential moved there. In site 1, the voltage-gated sodium channels failed to open despite electrical stimulation from site 2 because they were temporarily inactivated after the action potential was generated. In site 2, voltage-gated potassium channels opened, restoring the resting membrane potential after the passage of the action potential.
We already know that the outgrowths of neurons can be covered with neuroglial cells, or myelin sheath, wrapped around them. It is she who makes the axon look like an elongated chain of sausages; its formation and functions were discussed in. The transmission of the excitation potential along the "bare", not covered with myelinated cells, nerve fibers differs from its passage along the myelinated axon. When compared, the significant advantages provided by neuroglial cells wrapped around the axon are noticeable:

1 . Profitability. The axon membrane under myelin does not have channels for the passage of ions that create an action potential. Channels exist only in narrow spaces between glial cells, which are called interceptions of Ranvier(Ranvier's constriction), and only in these narrow sections of the axon does an action potential arise. The concentration of ion channels in these intercepts is 100 times higher than in the membranes of unmyelinated fibers, however, in order to ensure their operation, much less energy is required compared to how much energy would be required to provide ion channels located over the entire surface. naked axon. In addition, this economy allows the axon to “recharge” faster for the next action potential, so myelinated fibers are able to transmit signals of a higher frequency.
2
. Speed . On the "bare" axon, the signal is transmitted at each of its points. Ion channels open at every point on its surface, one after the other, which is reminiscent of careful, scrupulous reading into the text.
In a myelinated axon, an action potential that arose in one node of Ranvier reaches the neighboring nodes with its electric field, which leads to the emergence of new action potentials in them. That is, the excitation passes abruptly, from one interception to another. In case of damage to one intercept of Ranvier, the action potential excites the next interception after the damaged one, and, if necessary, even the third, fourth and fifth in a row. The spread of the electric field over this length is possible due to the electrical insulation created by the myelin sleeves: it reduces the scattering of the electric field, which is inevitable for unmyelinated fibers.
Thus, the rate of propagation of the action potential along myelinated fibers is much higher compared to unmyelinated ones. After all, in the same period of time, the action potential on the “bare” axon will excite only the area located directly next to it, and the action potential on the myelinated fiber will jump over a distance equal to the distance between one or more nodes of Ranvier:


The range of action potential transmission rates in various myelinated fibers is very wide: from a few meters per second to the "world record" set by the shrimp axon: it conducts excitation at a speed exceeding 200 m/s.
To imagine how effectively the speed of action potential conduction due to the myelin sheath can be increased, it is enough to compare the speed of its propagation along unmyelinated and myelinated fibers of human neurons. If we take the speed of impulse conduction in equally thin fibers, then in an unmyelinated fiber it will be approximately 1 m/s, and in a myelinated fiber 15-20 m/s. In thick human axons covered with a myelin sheath, the speed of conduction can reach 120 m/s.
So let's sum it up: the transmission of an electrical impulse through the body and processes of a neuron is carried out by passing ions through the cell membrane.

1 . first, a resting potential is created across the cell membrane. It is formed positively charged sodium ions ( Na+) outside the cell and negatively charged proteins inside the cell. The particles that create charges are located very close to the cell membrane, because they have different signs and are attracted to each other. Therefore, the resting potential exists only near the membrane; in general, the environment inside the neuron and outside it is electrically neutral. In addition to Na + and negatively charged proteins, they are involved in the creation and maintenance of an electric charge. positively charged ions ( K + ) and negatively charged chloride ions ( Cl-) also close to the membrane. The resting potential is approximately -70 mV (hereinafter, when indicating the numerical value of the membrane potentials, the word "approximately" will be omitted).
2 . when a neuron receives signals from other neurons through synapses, under their influence, on the membrane of the body of the neuron, postsynaptic potentials, that is, changes in the magnitude of electric charges on the inner and outer surfaces of the membrane. Under the influence of these signals, the internal negative charge and the external positive charge can increase or decrease by different amounts. And the neuron acts as a calculator summing up these effects.

3 . the result of the summation of postsynaptic potentials may be the continuation of the neuron in a state of rest. But if the sum reaches the value called excitation threshold, then in the axon hillock (thickening of the neuron body at the base of the axon) action potential.
4 . play an important role in the creation of the action potential cell membrane. It regulates the passage of ions through it (all of the above, except for negatively charged proteins that cannot leave the cell), opening and closing specialized channels that can conduct only certain ions.
5 . when the membrane of a neuron passes the threshold of excitation, it opens voltage-gated sodium channels. The obstacle that separated the positively charged Na + from the negatively charged proteins that attracted them disappears, and Na + rush into the cell. Thus, the external positive charge is transferred inside the cell, and the negative charge created by Cl - ions remains outside. There is a change in the membrane potential from a resting potential of -70 mV to +40 mV, and this is called action potential. It is important to understand that this change in potential occurs only in the area of the membrane where voltage-gated sodium channels have opened.
6 . At the moment the membrane reaches a potential of 40 mV, voltage-gated sodium channels close and open in the same area. voltage-gated potassium channels. Outside at this moment there is a negative charge, and inside - a positive one, created by the Na + ions that have entered there and the K + ions already there in large numbers. Potassium ions are pushed out into the environment surrounding the neuron through the opened channels, which sharply reduces the internal charge of the membrane and increases the external one. Charges of the same magnitude are created on both sides of the membrane, which would normally maintain the resting potential. But this state of the cell differs from the resting potential in that the potassium and sodium ions are not in their places: Na + is inside, and K + is outside. Again, everything described concerns only one place on the membrane, in which the action potential has just ceased to exist.
7 . To move the positively charged Na + and K + ions along the sides of the membrane needed to maintain the resting potential, it is used sodium-potassium pump. And this action also affects a limited area of the membrane: the one where the voltage-gated potassium channels have just closed.
One cycle of the sodium-potassium pump is that it captures three Na + ions from inside the cell, moves them outward, where it captures two K + ions and moves them inside. That is, the channel exchanges three Na + ions for two K + ions, and Na + can only move outward, and K + only inside the cell. Thus, the initial concentration of sodium on the outside of the membrane and potassium on the inside is restored. The sodium-potassium pump removes three positively charged ions from the cell, but only two enters it. Consequently, with each of its cycles, the internal positive charge of the membrane decreases, while the external one increases. In addition, the effluent potassium ions carry away their positive charges from the inside to the outside. The question arises: if the membrane potential was already equal to the resting potential after the entry of potassium ions into the cell, why lower it even more? The answer is that in addition to the ions mentioned here, there are others whose participation in the formation of the resting potential is less significant, and it is to compensate for their influence that such an equalization of the membrane potential is necessary. The result of this process is the achievement of the resting membrane potential: the charge of the inner side of the membrane is negative, the outer side is positive, the potential difference is -70 mV.
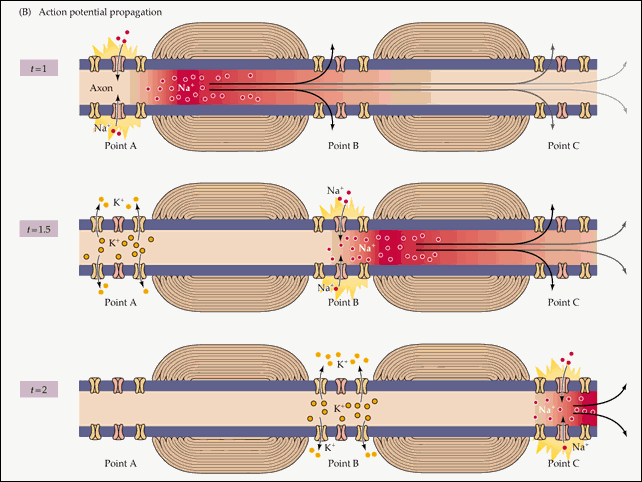
8 . So, in that place of the cell membrane, where only three steps ago there was an action potential, the resting potential again reigns. But the action potential did not pass without a trace: its existence (described in paragraph 5) created an electric field that excites parts of the membrane adjacent to it. On the one hand (towards the end of the axon), this excitation caused the opening of voltage-dependent sodium channels, triggering an action potential already in the new area. Which in turn generates an electric field that triggers the next action potential, and this cycle is repeated along the entire length of the axon, from its base to the ends connected to the next cells in the nerve chain.
On the other side of the action potential, closer to the body of the neuron, despite the influence of the electric field, the action potential does not arise. Because the voltage-gated sodium channels in this area have recently been opened, and for some time they are inactivated. It is this pause in their work that ensures that the action potential propagates in only one direction.
9 . Any signal, whether it is the transmission of information from a sensory neuron to the brain, or a brain command going to a motor neuron, is transmitted action potentials of the same size. The result of an action potential (the appearance of odor perception due to the work of sensory neurons, or the movement of the body closer to its source as a result of the coordinated work of motor neurons) depends solely on which nerve circuit the neuron is included in, this potential transmitting. To transmit the intensity of the signal (the strength of the smell or the magnitude of muscle contraction), the frequency with which action potentials are transmitted through the neuron is used. A weak signal is transmitted by rare, and a strong signal by frequent action potentials..
10 . the myelin sheath around the processes of the neuron increases the rate of propagation of the action potential due to the fact that the signal is transmitted over a long distance, equal to the width of one neuroglia cell wrapped around the process, and not to the area directly adjacent to the excited area, as occurs in unmyelinated fibers.
